���۷�������
0574-62101882


NMNȫ���������������ᣬ��һ����Ȼ���ڵ�������Ժ����ᣬNMN��2�ֲ����������ʽ:���ͦ¡����칹����NMN�Ļ�����ʽ��NMN��Ҫָ��-�������������ᣬ������Ϊ334.221 g/mol��
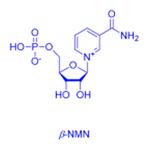
ͼ����Ч�Ħ�-NMN�Ļ�ѧ�ṹʽ
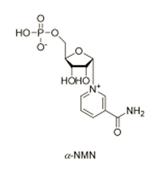
ͼ����Ч�Ħ�-NMN�Ļ�ѧ�ṹʽ
������������ά����B3�����NMN����ά����B�������ﷶ�룬��㷺�����������������Ӧ�������ߡ���лϢϢ��ء�
������������Ƴɵ�NMN��ҪΪ��-NMN��
1. NMN��ʲô���ü���ԭ��
NMN��NAD+��ǰ�壬�书��Ҳ��Ҫͨ��NAD+���֣��������Ҫ����һ��NAD+��
NAD+������øI��ȫ��������������˫�����ᣬ���㷺�ֲ������������ϸ���ڣ�������ǧ���������Ӧ���������ڱز����ٵĸ�ø��
NAD+�������ķ�Ӧ��Ҫ�����¼��֣�������DNA����PARPs�鵼����SIRTs���ס�NADP(H)�ϳɡ�
˥�Ϲ�����NAD+���½�����Ϊ�ǵ��¼����Ͳм�����Ҫԭ��������������ɥʧ����֪���˶������ϰ�������ȱ�ݣ�����������֢��Ӧʧ�����µĹؽ��ס���л�ϰ�����Ѫ�ܼ�����
��ˣ�����NMN���������NAD+�������Ӷ��ӻ������ơ���ֹ˥����صĶ��ֱ��ͣ��������յ��Ĵ�л���ҡ����꼲���ȡ����þ����У�

NMN/NAD��Ҫ����о�
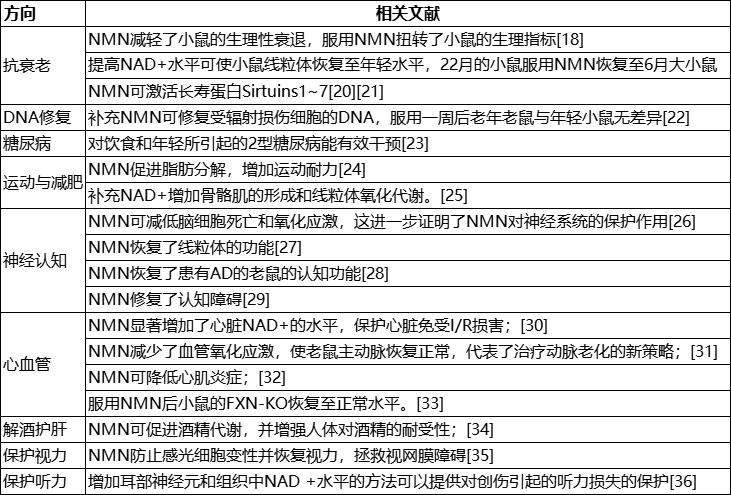
����о�ժҪ
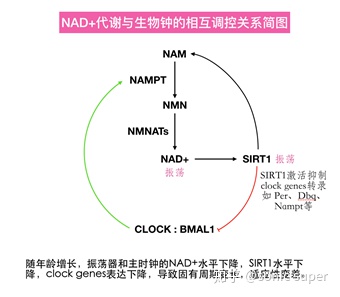
1.1NAD+����ҹ����
NAD+������������øSIRT1ͨ�����ӵ���NAD+����;����ø������·����ҹ����ת¼-���뷴����·����Ϊ��ҹ�������л֮���������
NAD+������������ͨ��SIRT1ʵ�ֵġ�SIRT1��BMAL1��PER2ȥ�������������CLOCK�����������������ģ�����SIRT1������CLOCK- BMAL1�鵼��clock genes��ת¼����ˣ�NAD+ͨ������ˮƽӰ��SIRT1ȥ���������ԣ��Ӷ�������Ӱ�����NAMPT���ڵ�һϵ����������ص��ı��
�����ӵ��ںͺܶ༲����أ�������������˯���ϰ��������������ܶಡ�����̶������������Ҵ������������ҿ����������Ŵ���Ҳ������Դ�ڻ������ܶ���֮����������������������ά�ֽ�����������Ҫ���á�
1.2NAD+����ϵͳ
Sirtuins��һ�����������������ʶ�������(NAD+)��������ø����ͳ����Ϊ���벸�鶯����������ƺ�˥���йء���Щ������˥�Ϲ����ж�ά����Ԫ�Ľ���Ҳ������Ҫ���á�
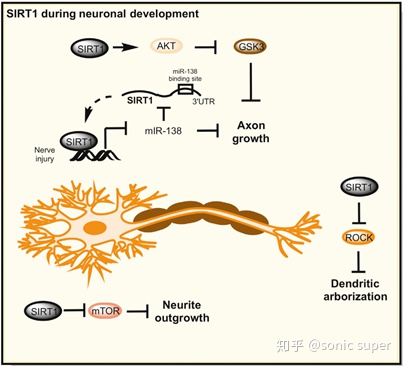
�����������У� SIRT1�ڽṹ��������Ҫ���ã�ͨ��Akt-GSK3ͨ·�ٽ�������������ͻ��������֦״��֧��ͻ���ķ�����ͻ��ǿ�ȵĵ��ڶԼ�����γ�������Ҫ����sirtuins���ײ����������������˺�����һ����������Ҫ�������á�SIRT1�ں����������������������ʽ���ڣ��ø���������ܵ���microRNA-134��ת¼����YY1��microRNA-134�ķֲ������������ԣ��ܵ���cAMP��Ӧ��ϵ���(CREB)����Դ����Ӫ������(BDNF)�ı�������ͻ�����γɺͳ��ڵ���ǿ������Ҫ��
������������չ�У�SIRT1�ڰ����ĺ�Ĭ�ϲ�������ɭ�ϲ����˶���Ԫ���ȶ����������Լ����з��ӱ������ã���Щ����������SIRT1�ڴ�л����Ӧ���ͻ������ȶ��Է���Ĺ����йء�����SIRT1��ҩ�����Ϊ������Щ�����ṩһ����ϣ���ķ�����
sonic super��NMN�ˡ������������²�֮��˭������zhuanlan.zhihu.com/p/74116762
1.3NAD+�방֢
����NAD+ˮƽ���ư�֢���о���ʾ����NMNAT3�����������������NAD+ˮƽ�����ƽ���ĸϸ����ϸ�����������ڲ���NA��NAM������SCIDС������������Ͷ���������ת�ơ�
��ԭ���У�������NAD+��ٽ�����������������ǽͽ⣬������ϸ��ϲ����Warburg��л�������������ữ���������ǽͽ�İ�ϸ��������л���ԣ�������NAD+Ҳ������SIRT1��SIRT6�Ļ��ԣ����߶�ͨ���µ���-catenin�źš��µ��ǽͽ�����������
�������Ҳ����ì���뵣�ǣ�NAD+�ٽ�DNA����Ѫ�����ɣ��п��ܰ�����ϸ�����������еĶ�Ұ����С��ij����о�δ���ṩ��ʹ�������ӵ��κ�֤�ݣ�������������NAD+ˮƽ������PARPs��DNA���˵��������ͣ���ϸ��/��֯�Ի���ҩ��������Խ����ӡ��ڱ���֢ģ���н�һ������NAD+�������Ч�����dz���Ҫ��
1.4NAD+����
��֪NAD+�ź�ͨ·�е�ø���Ա��������֬���ѻ�����ά�����ȵ��صֿ���Ӱ�죬��Щ����֬���μ����ķ����йء�
NAMPT�ڸ�֬��ʳ�յ�֬���η�����չ�Ĺ�������ؼ��������ã�����NAMPT��ʹ��֬��ʳ��ɵĸ�֬�����Ը����أ�������NAMPT�������Ƹ�֬�ʻ��ۣ����ֵ���������ͨ��������NAMPT������NAD+������SIRT1������SREBP1��ȥ��������SREBP1���Խ��͡�FASN��ACC�����ϵ��������ġ�
SIRT1�������ΰе�PGC-1a��PSK9��SREBP1ά�������幦�ܡ����̴�ת�˺�֬������̬��SIRT2ͨ��ȥ����������ϩ����ͪ���ȼ�ø��������������SIRT3����OXPHOS��֬����������ͪ���ɺͿ�����Ӧ����SIRT6������������
������Щͨ·�ڸ����е���Ҫ�ԣ�ά��NAD+ˮƽ����ά���������ù��ܱز����١���������£����ڷ��ֺ�˥�ϣ�NAMPTˮƽ�½���CD38ˮƽ���ߣ����µ�����ʱ����̬NAD+ˮƽ�½�2����
��NAD+ˮƽ��ߵ�����ˮƽ��Ԥ�������Ʒ��֡��ƾ���֬���Ը���NASH������Ч������ͬʱ���ܸ�����������̬�������幦���ϰ������Ƹ���Ľ�������ǿ�����������������������ܸζ�����
1.5NAD+��������
����������NAD+ˮƽ�Ľ��ͺ�sirtuin���Ե���Ӧ�����ںܴ�̶����������ܺ�˳Ӧ���������½���ԭ��
��ͨ��NAD+���伤��SIRT1��SIRT3���������յ�����ϵĤϸ���ʴ���NMN����С����SIRT1�����Եķ�ʽ����˳���յ��ļ���������(AKI)��
��5-��������-4-�Ȱ����տɴ̼�AMPK���ԣ�����NAD+ˮƽ������sirt3�����ķ�ʽ����˳���յ���AKI��
��С��NAM�ɴ̼����ౣ��ǰ������PGE2�ķ��ڣ�����ȱѪ�������ܣ�NAMҲ��ͨ���̼�NAD+�ϳ�����˳���յ���AKI ��
1.6NAD+�������
�������Ұ����С����ȣ�С��ļ���ή������֢��־���Լ��ȵ����źź��ȵ��ش̼���������ȡ�����½�����NAD+ǰ����������С����������Ƽ���ܡ�
��NMN (500 mg/kg/day ip .����7��)��������С����ͨ�����������幦�ܡ�����ATP���ɡ�������֢�����ǽͽ�II�ͼ���ת��Ϊ������ά�ͼ��⣬��ת��������ص��к��仯��
1.7NAD+�������
NAD+ˮƽ����������ܺ����˺�Ļָ�������Ҫ��������NAD+�������źŵ����У�SIRT3�ƺ�������Ҫ�ģ�
��SIRT3�ó�С���OXPHOSø�߶���������ATP���٣��������������߶����У�����������������ͨ�Թ��ɿĵ�������CypD�ļ��
��SIRT3-KO����13���´�ʱ�ͻ������ά�����ļ��ʺ��������������������һ���Ӿ磬��NMN���ƿ�����ת�����½���
��������ȱѪǰ30����(500 mg/kg, i.p.)���ٹ�עǰ���ٹ�ע�ڼ���ظ���ҩ��ʹ��NAMPT�������NMN���ƶ���������ֹѹ�����غ�ȱѪ-�ٹ�ע���ˣ�ʹ�����������44%���ҡ�
��ʹ��NAD +ǰ������Ҳ���������MDX�ļ���С�������ܡ�
��NAD+ǰ�������ȱ���յ�������˥��С��ģ�͵������塢����ܡ�
��NAD+ǰ����������ͨ������SIRT3�������ָ������ϣ�Ϲ���ʧ��(FRDA)�ļ���С��ģ�͵�����ܵ���������ˮƽ��
1.8 NAD+��Ѫ����Ƥϸ��
��Ƥϸ����EC��˥����һ���ṹ���ܸı�IJ����������̣�����Ѫ������ʧ������Ƥͨ�����ӡ�����Ӳ����Ѫ�����ɺ�Ѫ��������EC�������������ɼ��ٵȡ�
ϸ������ʧ��������Ӧ�������źŸı䡢������Ѫ֢��Ѫ����֢��EC˥�Ϻ�Ѫ�ܼ����ķ�������չ������ء������쳣�ķ���ͨ·����ЩDZ�ڵIJ��������仯�йأ�����SIRT1��Klotho������άϸ����������-21������Ѫ�ܽ�����-ȩ��ͪϵͳ�ļ��
��ΪSIRTs��Ѫ��˥�ϵĹ�ϵ��NAD+ǰ��NMN�IJ��䲢���Ѿ���һЩ�о������ֳ�Ч����
��NMN��������С��(8����ÿ���ҩ300 mg/kg)�ɻָ���������Ƥ����������(��Ƥ���ܵ�һ�ֲ�������)��ͬʱ�����������������ٺ͵��Զ����նȡ�
��NMN (500 mg/kg/�죬ˮ�ͷ�������28��)��С�������ȡ��������Ч��ͨ���ٽ�sirt1-������ëϸѪ���ܶȵ����ӣ�����������С���ѪҺ������������
��NMNͨ����������С�������յ���Ѫ����Ƥ�����ϰ��Լ���Ѫ����ϣ�NVC����Ӧ�������������С����֪������NMN������������Ѫ����Ƥϸ����������ROS���ָ�NAD+��������������
��Ѫ����Ƥ������NAD+ˮƽ�����ܳ�Ϊһ�����������˵Ļ����DZ���Ʒ�������������Ѫ�����ٶ�������չ�ļ����磺ȱѪ-�ٹ�ע���ˡ��˿����ϻ������ι����ϰ��ͼ��⼡���ȡ�
1.9 NAD+���л�ϰ�
NMN��֬����л���Ǵ�л���ҵ��µķ��֡�����������ֳ���ƶ��и������ã������ܹ����Ʒ���ĸ�Դ��Ժ����ֳ�IJ���Ӱ�졣

NAD+ǰ���ڶ���ʵ���жԴ�л�ϰ�����������
2. NMN��NAD+
NAD+����ô�����ã�����Ӧ����β���������Ϊ��ѡ��NMN��

��Ȼ�Ƚ��ܺϳ�NAD+����Ҫ;����
NAD+�ĺϳɸ��ݲ�ͬ�ϳ�ԭ�Ϸ�Ϊ����;������ͷ�ϳ�;����Preiss-handler;����
a) ��ͷ�ϳ�;������ɫ���ᣨTrp��ת��Ϊ����ᣨQA����Ȼ��ͨ�������-������ǻ�ת��ø��QPRT��ת��ΪNAMN��NAMNת��ΪNAAD�������վ���NAD+�ϳ�ø��NADS��������NAD+��
b) P-H�ϳ�;�����ֽ�NA����;���������ᣨNA����NAPRT��NMNAT��NADS��NAD�ϳ�ø���ϳ�NAD+��
c) ���Ⱥϳ�;�����ֽ�NR����;���������������ǣ�NR������������NAM����NRK�����������ռ�ø����NAMPT��NMNAT�ϳ��������������ᣨNMN����NMN��NMNAT1-3ø�ϳ�NAD+��
����������գ�NAR��ͨ��NRK��Ҳ�ܲ���NAMN�����1�е�ø�ٷ�Ӧ�ϳ�NAD+��
PNP�����ʺ������ữø��NRK�����������ռ�ø��QPRT��������������ת��øNAPRT�������������ת��ø��NAMPT���������������ת��ø��NMNAT����������������������ת��ø
��NAD+�����ںϳɾ�����֯�����ԣ�
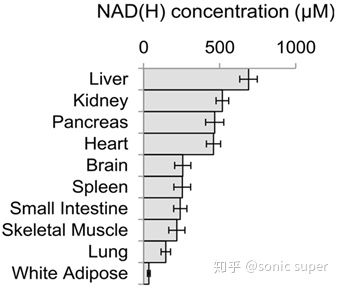
���鶯�������֯����NAD(H)Ũ��
���鶯�����֯���ٶ�NAD+ԭ�ϵ�ƫ���ԣ���Ҫ�����DZ����NAD�ϳ�ø��֯�������йء�
����ʹ��ɫ�����ͷ�ϳ�NAD+���ںϳ�-ʹ��NAD+ѭ�������з��ڴ���NAM(������)����Щ������������ѭ��������������֯��ȡ���á�
��Դ�����ᣨNA����ѪҺ-��֯���ٵ���������ٺ������������֯���ٲ�������ѪҺ�е�NA�ϳ�NAD��
���������������֯��Ϊ����ѭ��ϵͳ���͵�NAM�ϳ�NAD������������ȡNAM�ϳ�NAD+Ч����͵���֯��С����Ƣ��������NAM�ϳ�NAD+Ч����ߵ���֯��

���鶯�������֯����NAD(H)���ֺϳ�ø�ı���
����һ�����º������Ȼ���䣺NMNĿǰ�Dz���NAD+����ѷ�ʽ��
2.2 NMN�����岻ͬ�������в�ͬ����
NMN�ĺϳ�ø������øҲ������֯�����ԣ�
NMN��ȫ����֯���������й㷺�ķֲ����Ҵ���̥����ʱ�ھʹ����ڶ���ϸ���С�
��NAD+ǰ���ڸ�����֯��ϸ���ڵĴ�л������ֲ�֪֮���٣����֮���˽�϶����NMN�ĺϳ�øNAMPT��NRKs���Լ�NMN����øNMNATs�ı��
��1��NAMPT
NAMPT�������������ڣ�����֯�����ˮƽ���ڽϴ���졣���Ժ����࣬NAMPT�����IJ���;���Dz�NAD+����ѡģʽ�����ڹ�������NRK�����IJ���;���Dz�NAD+����ѡģʽ��
��2��NMNATs��NMN����ø��
С����֯��л�ױ�����NMNAT���͵Ļ���Զ����NAMPT���ҳ�ѪҺ�⣬�������֯��NMNAT���͵Ļ��Բ������ơ�
��3��NRKs
NRK���͵ı����������NRK1�������ڣ���NRK2��Ҫ�����ڹ������С����һ�µ��ǣ�����NR�����������NAD +ˮƽ���ӣ����ڴ��Ի��ɫ֬����֯��Ч����
�ڷ�NMN��NAD+�Ĵٽ���
��Ȼ������Ѫ���м������ṹ��NMN���ڷ�NMN���ܹ��ܿ죨15min����ߴ��ԡ�����С���NAD+ˮƽ��

���ࡢ���١���ɫ֬����֯NMN��NAD+ˮƽ
2.3 NMN��ν���ϸ��

NMN����ϸ���IJ�ͬ;��
NMN��ijЩϸ��������Ĥת���壬��ֱ�ӽ�NMNת��ϸ���ڣ�����NMN�����ֽ���ϸ���ķ�ʽ��
��ͨ��ת����ֱ�ӽ���ϸ������2019�����nature metabolismһƪ����֤ʵ�˸��뷨�����·���С��С������NMN������ת������ڣ�����Slc12a8������һ�ְ�����Ͷషת���壬��NMN�кܸߵ�ѡ���ԣ�����ת�˺�NMN�ṹ��Ϊ����NaMN��
��ͨ��ϸ��Ĥ�����CD73ȥ���ữΪNR��ͨ��ƽ�����ת�˵���ENTs������ϸ���ڣ������ͨ��ϸ���ʵ�NRKø��ΪNMN�����������屻���ã���������NRK����
NAM����NMN��ǰ�壬����NAD+��NADase��������;��CD38ˮ���IJ�����NAD+�ĺϳɡ����á�������һ���漰���ڰ����NMN/NR��NAD+��NAM��NMN��ѭ����
3 NMN����Դ����Щ��
3.1 ��Ȼʳ��
NMN���ճ�ʳ���зֲ��Ϲ㣬�߲��绨Ҭ�ˣ�0.25�C1.12mg NMN/100g���ʹ�ײˣ�0.0�C0.90 mg NMN/100 g����ˮ�������棨0.36�C1.60 mg NMN/100 g������������0.26�C0.30 mg NMN/100 g������������ţ�⣨0.06�C0.42 mg NMN/100 g�������зḻ��NMN��

������ѧȨ����־Cell Metab. ��Ӱ������22.415���ܽ�����NMN��ʳ��
��Ȼʳ����Ҳ����NMN�������Dz��ǿ���ͨ���ճ�����ʳ�������أ�
��������ѧȨ����־Cell Metab. ��Ӱ������22.415���ܽ�����NMN��ʳ����Կ���������������ڳ���NMN����Ʒ��һ������Ч�ɷֺ���150mg����Ļ���Ҫ��ﵽ�ͳ�һ����-��������������/NMN����Ʒͬ����Ч����������Ҫ��16~64��ë����27~120����������19~83��ţ�����������������͵�ʳ�����ǾͲ������ˡ��� ����Ԥ�ƣ�����ͬ�ȼ���������²���û��ʡ����Ǯ�����ǵ�θ���ǻ��ȱ����������뵥���ճ���ʳ�������-��������������/NMN�����Ƿdz����ģ�������ʵ������Ŀǰ���æ�-��������������/NMN��ĿǰΨһ�IJ���;����
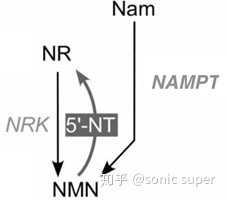
NMNҲ���Ծ���Դ�����ʺϳɣ�1������������1����5-������ǻ�-1-�����ᣨPRPP�����������������ת��ø��NAMPT��NAMPRT��������������1����NMN��1���ӽ����ᣨPPi������������������NMN��1�������������գ�NR�������������ռ�ø��NRK���������ữ����1����NMN��
3.2 �ϳ�NMN
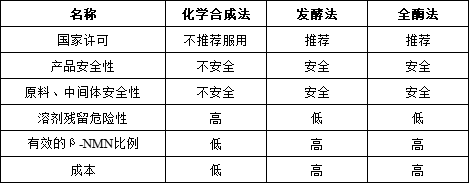
NMN�ϳɷ�����������ѧ�ϳɷ������ͷ���ø������
��ѧ�ϳɷ����ڸ���ԭ�ϼ۸�����ѣ�������·�̳����������ѡ��ɱ��ߡ������칹�������롢������ý������Ͷ����һ�ֻ����ȱ�㣬������Լ��NMN�Ĵ��ģ��������Ӧ�á�����Ŀǰ�й�������ʳƷԭ��ԭ���ϲ��Ƽ���ѧ�����Ʊ�������ԭ�ϣ���������ϳ�Ϊ���ѡ��
����ͷ�����ת���ʹ��͡�NMNŨ�ȹ��ͣ����·����Ѷȸߣ��ۺ�ԭ�ϡ�����ɱ���ܸߣ������˷��ͷ��ķ�չ��
����������ѡ��һ�ֻ����ø����ͬ�����������ϳɣ���Ϊ��ѡ�о��ķ���
��Ҧ����˹�������Ƽ�����˾
��ַ���㽭ʡ��������Ҧ���ÿ����������³��˺�·9��
���۷���������0574-62101882

�Ź��ں�

�ٷ��̳�
Copyright © 2021 ��Ҧ����˹�������Ƽ�����˾
