���۷�������
0574-62101882


Samir M. Parikh
��øNAD+��ɫ�����л����о���������С��ģ���У�ͨ������ø��;���е��м���Ľ��⣬�������������β���
�ݹ۵�����������չʷ������������С���Ӱ����߾������صĽ�ɫ����������������Ŵ����룬���������ᴫ���źŻ����������⣬������Ķ�������ʽ����Ϊ�����������ʶ������ᣨNAD+������֤ʵ���������ֹؼ���ϸ�����ܡ�
һ�������Ӵ��͵�һ�ֱ���Ϊ�������ϸ������ø�������У������õ�����Ч�ػ�ȡ��
������Ϊø�ĵ������ȥ������ø��Sirtuins�������ض���ϸ����Ϊ��
��Nature��һƪ�����У�Katsyuba ����1��ʾ����ϸ��ˮƽ�ϣ�NAD+�����û��ƣ���֤ʵ������NAD+���Ӱ�켲��������չ��
�����֮��ϸ���п��õ�NAD+ȡ���������ɺ�����֮���ƽ�⡣�����ද�����NAD+����Ҫ;����������������NAM������ѭ�������ַ���Ҫô����ȡ��Ҫô���ֽ�NAD+��øø���ͷţ�ͼ1������������������NAD+�ĺϳ�·�ߣ����������谱����ɫ���ᣨTrp���Ĵ�ͷ�ϳ�;��2��ͻ���ƻ��˸���ɫ����ת��ΪNAD+��ø������������ϵͳ�ķ�����ȫ3���Ӷ�֤ʵ�����ִ�ͷ�ϳ�;������Ҫ�ԡ�
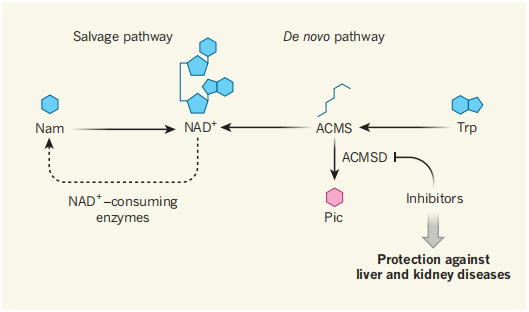
ͼ1��������NAD+������ϳ�
��øNAD+��ø��������������Nam����ͨ�����Ⱥϳ�;����NAD+ѭ��������Nam�IJ��Ⱥϳ�;������Ϊ��NAD+����Ҫ�ĺϳ�·�ߡ�����NAD+�Կ�ͨ�������ܶ�;���ϳɡ�����һ������ɫ����Ĵ�ͷ�ϳ�;����ɫ���ᾭ�������м��壬���ձ�ת��ΪNAD+�����а���ACMS����һ;�����Ա�ACMS����ø��ACMSD���жϣ�ACMSD���Խ�ACMS��Ϊ��़��ᣨPic����Katsyuba ������Ϊ��С����߳����ڣ�ͨ����ѧ�ֶ�����ACMSD��������NAD+��ˮƽ���Ӷ���Ч����С��������β�״����
Katsyuba���˿�ʼ�о���-����-��-�Ȼ�����-��-��ȩ��ACMS����һ���о����ٵIJ��ȶ���ɫ����;�����м��塣ACMS�����Է���ת��ΪNAD+;���е���һ���м��壬���ҿ��Ա�ACMS����ø��ACMSD����һϵ��ø���⡣��ˣ�����Ԥ�⣬�ڴ�ͷ�ϳ��У�ACMSD����NAD+�����ɡ��������߳滹��С��ACMSD�ڽ����϶��DZ��ص�4������һ�����˵ķ��֣���Ϊֱ��������߳滹û�б��Ͽ��Դ�ͷ�ϳ�NAD+��
���������ߣ��������߳��б���ACMSD��acsd-1���Ӷ������NAD+��ˮƽ��������֪������NAD+�����ӳ��������������߷��֣�������������棬���ڵ�acds-1�ı�����ȫ����ϡ����⣬����acsd-1�ı����������뿹˥����صķ��ӷ�Ӧ5,6����������ȥ������ø-2.1�Ļ��ԣ����������幦�ܣ������Ե�������Ӧ����Ӧ��
�������С�����ڣ�����������������ACMSD7��������о���������Щ��ɫ����������NAD+����;������Ҫ����8��Katsyuba���˷�������Acmsd����������С���ϸ��NAD+��ˮƽ����ǿ�����幦�ܡ���ˣ������߿�����ACMSD�Ļ�ѧ���Ƽ���������С�����ڣ���������Щ���Ƽ��Ƿ��ܸ�������˥����صļ�������ʳ��֬�����뼱�������ˡ�
�й�NAD+�ı������о����Ѿ�������9,10��Katsyuba���˵Ľ��֤ʵ������NAD+�ڼ��������е�DZ��������һЩģ���У�ʹ�����ǵ����Ƽ��������Ƽ������о������������NAD+�Ĵ�ͷ�ϳ�;�������Ӿ����Ը������NAD+ˮƽ��صĸ�������༲����Ȼ����Ҫ֤����һ�㣬����Ҫ֤��ACMSD���Ƽ������û���������NAD+����������Ļ��ƣ��������ᣨ��ACMSD�鵼��ACMs��������ģ��ĺĽߡ�����õ�֤ʵ����һ���ֽ�������һ���о������һ��11�����о���ɫ����ͨ·�з�����һ�ֲ�ͬ��ø���ŵͪ�������ǻ�ת��ø���Ǽ������������Եľ������ء�
�м�������ֵ������˼�������磬��ʲô����ѹ����ʹNAD+�Ķ�������ϳ�;���ı��������أ�Ϊʲô���漰�����鶯�����ڵĽⶾ���٣���ͷ�ϳ�;����Ϊ��Ծ? һ��ֵ��ע��Ŀ����ǣ������������������ٸ������ܵ���������ţ�����NAD+����ʵ�ϣ���Щ���ٰ�Nam����������������λҲ�����Խ��ͽ����뼲�������ټ�Ĵ�л��ϵ�������磬Ϊ�����Ըβ����ߵĴ��Ժ�����ܾ�������8��
Katsyuba���˿�����ACMSD���Ƽ���ʾ����NAD+���ӵ��ٴ�����DZ�����ӵ�һ��NAD+����Ϊ�������ľ������ص����Ѿ�������ȥ��20��12����������˥�ϵĸ����ԣ����ѽ��䶨�塣�й�����NAD+�������������ϵ������̫��ʱ�������ڲ��������ޡ��෴�����˥�ϵĶ�������˶Լ���Ӧ��Դ�����Ⱦ�����˻��������ĵֿ�����������ôNAD+���ٴ������ܱ�ø��ӿɲ����ԡ��������һ���о�Ӧ������һ����������һ���ڽ������������������Ⱥ�н��еĿڷ�Nam���Ƶ�����11�����������һ�������������������������ˣ����������������йأ���NAD+�����ӱ����˼��������ˣ���Ȼ��ֻ�dz����Ľ������������NAD+�������о�ǰ����
Ȼ�����ڷ�NAD+ǰ����ܲ���һ������NAD+����Ч����8����ˣ���Katsyuba����ͬ�¿�����ACMSD���Ƽ��������Ͻ����м�ֵ�ġ����ǵ���������д�ͷ�ϳ�;����ø�ĸ�������������IJ���Ҳ�������һ����Ȥ��˼·����֯�����Ե�����NAD+��������NAD+����Ч;����
��NAD+������DZ�ڵ������õ�;���Ƕ��ֶ����ģ��������13���������Լ���14�ʹ�л�ۺ���15��ʹ�ò�ͬ�ķ����������Ŵ�ѧ3�����仯ѧ8���Ƚ�ϵͳ����1���ٴ��о�11����һϵ���о�����ɫ����ͨ·��NAD+ˮƽ����Ҫ�����ߺ�ҽ������̽������Ҫ����
�ο����ף�
1.Katsyuba, E. et al. Nature https://doi.org/10.1038/ s41586-018-0645-6 (2018).
2.Krehl, W. A., Teply, L. J., Sarma, P. S. & Elvehjem, C. A. Science 101, 489�C490 (1945).
3.Shi, H. et al. N. Engl. J. Med. 377, 544�C552 (2017).
4.Fukoka, S.-I. et al. J. Biol. Chem. 277, 35162�C35167 (2002).
5.Mouchiroud, L. et al. Cell 154, 430�C441 (2013).
6.Gomes, A. P. et al. Cell 155, 1624�C1638 (2013).
7.Pucci, L., Perozzi, S., Cimadamore, F., Orsomando, G. & Raffaelli, N. FEBS J. 274, 827�C840 (2007).
8.Liu, L. et al. Cell Metab. 27, 1067�C1080 (2018).
9.Tran, M. T. et al. Nature 531, 528�C532 (2016).
10.Gariani, K. et al. Hepatology 63, 1190�C1204 (2016). 11.Poyan Mehr, A. et al. Nature Med. 24, 1351�C1359.
11.Poyan Mehr, A. et al. Nature Med. 24, 1351�C1359 (2018).
12.Lin, S.-J., Defossez, P.-A. & Guarente, L. Science 289, 2126�C2128 (2000).
13.Williams, P. A. et al. Science 355, 756�C760 (2017).
14.Wang, G. et al. Cell 158, 1324�C1334 (2014).
15.Cant��, C. et al. Nature 458, 1056�C1060 (2009).
��Ҧ����˹�������Ƽ�����˾
��ַ���㽭ʡ��������Ҧ���ÿ����������³��˺�·9��
���۷���������0574-62101882

�Ź��ں�

�ٷ��̳�
Copyright © 2021 ��Ҧ����˹�������Ƽ�����˾
